Природная яркость аквариумных тропических рыб поражала людей издавна: сначала выловленных из природных водоемов, а потом и наблюдаемых в аквариуме. Морские и пресноводные рыбы по разнообразию окраски могут поспорить с другими животными. В этой статье мне хотелось бы остановиться на функциях окраски и общих механизмах ее формирования и регуляции у морских и пресноводных рыб.


Сначала несколько слов о функциях окраски. «Это не очень интресно», — скажете вы. Преднерестовый окрас самцов большинства видов рыб, предупредительная и маскировочная окраска, и т.п. — все это давно известно. Одно «НО». Для чего нужна окраска, например, морским рыбам, обитающим на глубине 20 м и ниже? Ведь там невозможно увидеть весь цветовой спектр, которым так богаты тропические рыбы.
Значит, эволюционно сложилась другая, более ранняя, функция окраски. И эта функция — выделительная. Дело в том, что для того, чтобы вывести ядовитые вещества, образующиеся в ходе обмена веществ, их необходимо предворительно перевести в нерастворимую форму или полимеризовать. Такие полимерные соединения могут хорошо поглощать свет, становясь пигментами.
Гуанин, птерины и некоторые другие пигменты содержат много азота и эволюционно являются конечными продуктами метаболизма вредных веществ, откладывающихся в кожных покровах и со временем сшелушивающихся. Известный биолог А.Е. Микулин [А.Е. Микулин «Функциональное значение пигментов и пигментации в онтогенезе рыб», Изд. ВНИРО, М., 2000] отмечает еще и важную антиоксидантную функцию пигментов. Мозг многих рыб снаружи выстлан меланофорами, которые регулируют подачу световой энергии. Было также показано, что каротиноиды в развивающейся икре могут задерживать кислород, что способствует быстрому ее развитию. Кроме того, некоторые пигменты спасают икру от переохлаждения, но это уже тема другой статьи.
Яркость окраски у рыб зависит от количества клеток - хроматофоров и пигментов, которые в них содержатся. Цветовой потенциал формируется посредством правильного питания, которое позволяет правильно развиваться хроматофорам, а именно делиться и запасать пигменты, и генетической предрасположенности к такому развитию.
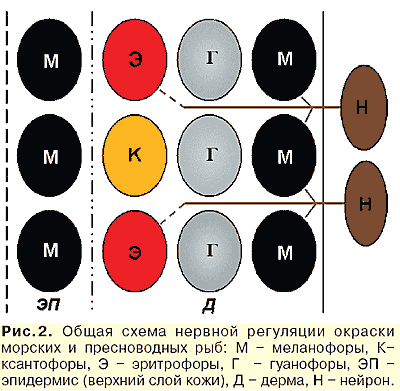
Хроматофоры бывают четырех видов — меланофоры (содержат пигмент меланин) придают черную окраску, ксантофоры (пигмент лютеин) — желтые и оранжевые тона, эритрофоры (пигмент астаксантин) — красный цвет и гуанофоры (кристаллы гуанина) — металлический блеск. В кожных покровах они располагаются в том же порядке от верхних слоев кожи (эпидермиса) к нижним (собственно дермы). При этом меланофоры могут располагаться как в эпидермисе, набирая цвет по аналогии с загаром и светлея от сшелушивания, так и в глубинных слоях дермы. Хроматофоры располагаются в разных слоях кожи, поэтому цвета, определяемые пигментами, накладываются друг на друга и образуют разные оттенки. Соответственно, от наличия или отсутствия этих клеток (и пигментов, содержащихся в них) зависит окраска данного участка кожи рыбы. Вспомните, как в детстве вы рисовали красками: синий и красный — дает фиолетовый, синий и желтый — зеленый и т.д. Смесь трех-четырех цветов дает еще более интересные оттенки.
Так, например, голубому неону (Parachdrodon innesi), гуанофоры придают блестящую окраску. Мраморные гурами (Trichogaster trichopterus "Cosby") имеют меланофоры, которые позволяют ему быстро изменять окраску от почти черной до светло-голубой. Каким образом? К ним подходят нервные окончания, которые влияют на эти клетки и заставляют их перераспределять внутри себя меланин таким образом, чтобы пигмент был заметен или нет.
К гуанофорам нервные окончания не подходят. Есть научные работы, в которых показано, что к эритрофорам тоже подходят нервные окончания (рис.2).
Теперь поговорим о накапливании пигментов. Меланин синтезируется из аминокислоты тирозина. Кстати, промежуточным веществом такого синтеза является предшественник адреналина и дофамина («гормона радости»). Эритрофоры содержат преимущественно астаксантин, а ксантофоры — лютеин. Предшественниками эритрофоров и ксантофоров у молоди рыб являются птерины, цветность которых со временем замещается каротиноидами, полученными из пищи. Гуанофоры содержат кристаллы гуанина (рис. 3). Каков же механизм изменения окраски?
Существует три основных типа регуляции окраски: нервная и гуморальная регуляции, а также изменение содержания внутриклеточного и внеклеточного кальция.
К меланофорам подходят нервные окончания. Нервные импульсы отменяют или разрешают рыбе окраситься в черный (а на самом деле и некоторый другой) цвет. Если пигментные зерна расположены по всему хроматофору, то происходит экспансия пигмента (усиление окраски), а если все пигменты собраны в центральной части клетки — аггрегация пигмента. Для остальных хроматофоров цветность может регулироваться гормонами (гуморальная регуляция): агрегацию пигмента вызывают, например, витамин С, пирокатехин, тимол и вератрин, экспансию — никотин, атропин, ряд жирных кислот и ацетилхолин. Интересно, что адреналин может вызывать одновременно агрегацию пигмента в меланофорах и экспансию уже другого пигмента в гуанофорах. Пролактин вызывает экспансию пигментов в эритрофорах и ксантафорах. Примеров можно привести очень много. Но самое интересное, на мой взгляд, — это регуляция типа окраски посредством изменения содержания внутриклеточного и внеклеточного кальция. Во всех типах хроматофоров, в которых содержатся ка-ротиноиды, направленный вход ионов кальция в эти клетки ведет за собой агрегацию этих пигментов. И наоборот: если кальций выходит из клетки, то экспансия пигмента усиливается. При этом кальций связывается с молекулами каротиноидов и окраска меняется. Типичный пример входа кальция — воспаление, как инфекционное, так и нет. Назовем это более понятным словом — болезнь.
Теперь стало понятно, почему больная рыба слабо окрашена (более бледная). Не потому что она теряет пигменты, а из-за связи ионов кальция с пигментами.
Конечно, я совсем упростил схему. Существуют и другие причины. Например, цитоскелет клетки — это как зонтик и спицы: в сложенном состоянии видна лишь часть зонта с определенным цветом (если конечно зонт не мужской  ), раскрытом — все цвета зонтика. Так и в клетке — от степени разборки цитоскелета (спиц) зависит степень показа всех его пигментных участков. Для гуанофоров — это влияние на поворот кристалла (см. выше). Важно отметить, что гуанин может превращаться в птерины. Из ксантофоров в процессе онтогенеза происходят эритрофоры.
), раскрытом — все цвета зонтика. Так и в клетке — от степени разборки цитоскелета (спиц) зависит степень показа всех его пигментных участков. Для гуанофоров — это влияние на поворот кристалла (см. выше). Важно отметить, что гуанин может превращаться в птерины. Из ксантофоров в процессе онтогенеза происходят эритрофоры.
Таким образом, можно говорить о множественной функции окраски морских и пресноводных рыб, не ограничивающейся зрительными образами и эволюционно обладающей как выделительной функцией, так и антиоксидантной и другими видами протективной активновти.


