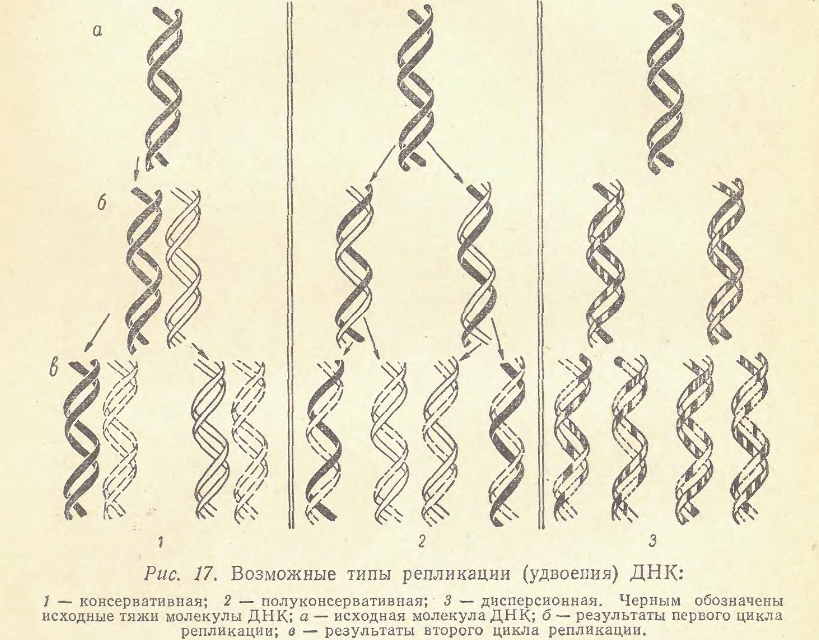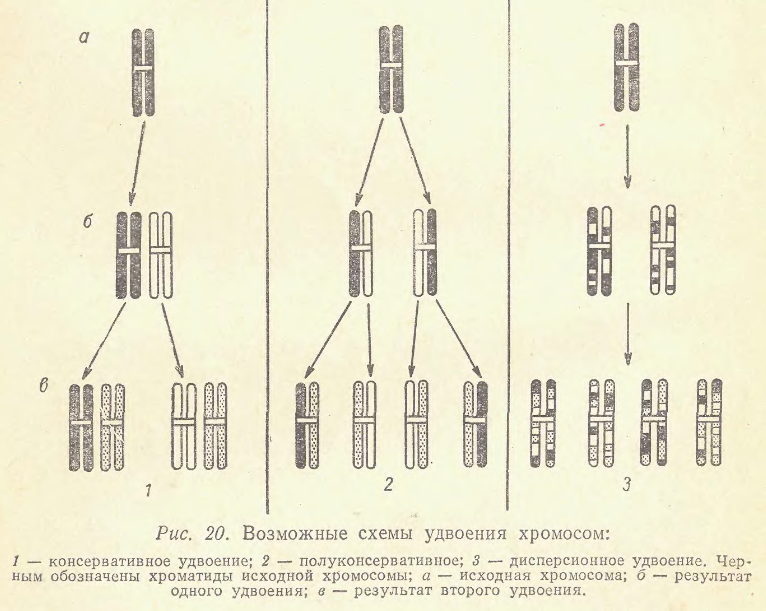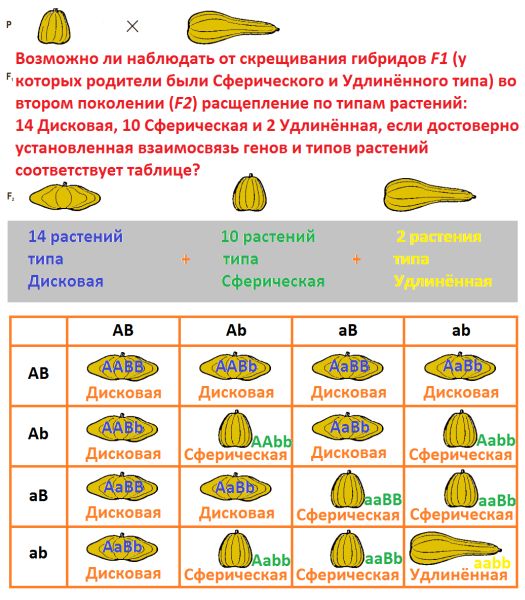
Продолжим...
(цит. по Тарантул В.З. Геном человека: Энциклопедия, написанная четырьмя буквами. М.: Языки славянской культуры, 2003. — 392 с. Версия online)
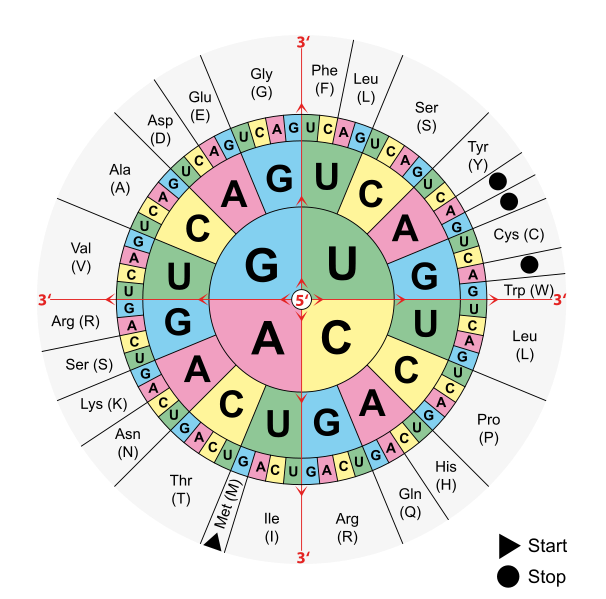
После получения всех этих новых многочисленных сведений об особенностях организации и эволюции генов у человека (сходным образом обстоит дело и у других высших организмов) стало ясно, что дать строгое определение того, что же такое собой представляет ген, довольно сложно. Но даже если бы такое определение сегодня имелось, его, как и всякое другое, не следовало бы считать незыблемым. Не зря говорил Спиноза: «Всякое определение есть ограничение». Однако в целом базовое представление о гене осталось в силе. Ген — это физическая (определенный участок ДНК) и функциональная (кодирует белок или РНК) единица наследственности.
...
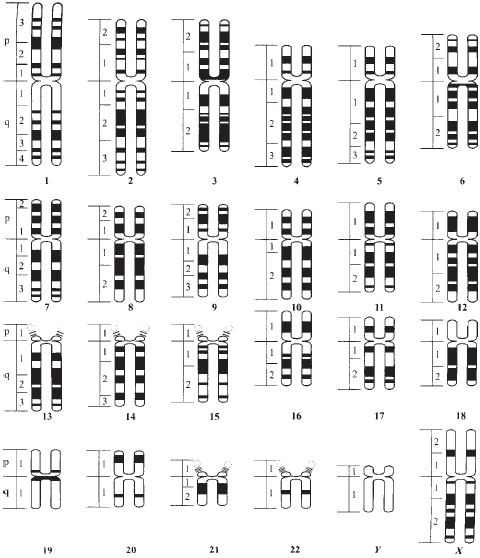
Весьма интересные результаты были получены, в частности, при сравнении распределения по хромосомам сходных последовательностей ДНК человека и других животных. Приведем лишь один пример. Как уже указывалось, между геномами человека и мыши имеется большое сходство. На рис. 37 на цветной вклейке изображено расположение в разных хромосомах мыши сходных сегментов отдельных хромосом человека. Глядя на этот рисунок, мы можем увидеть, что участки одних и тех же хромосом человека распределены во множестве хромосом мыши. Это справедливо и наоборот. А что это значит? Это говорит нам о тех путях, по которым шла эволюция млекопитающих (ведь мышь и человек млекопитающие). Тщательно проанализировав картину, изображенную на рис. 37, ученые установили, что на границах разных участков ДНК мыши, которые обнаруживаются в составе ДНК человека, содержатся различные подвижные генетические элементы, тандемные повторы и другие «горячие точки», по которым, вероятно, и шла перестройка (рекомбинация) в ходе многовекового процесса эволюции животных организмов.
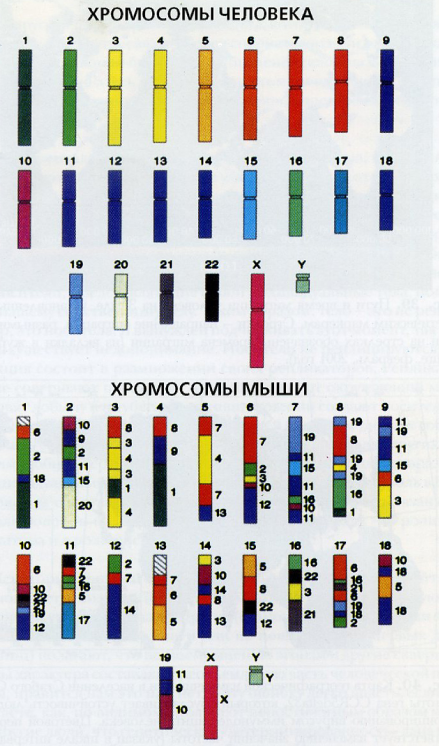
Рис. 37. Генетическое сходство (гомология) хромосом человека и мыши. Разными цветами и номерами на хромосомах мыши отмечены нуклеотидные последовательности человеческих хромосом, содержащие сходные сегмент.
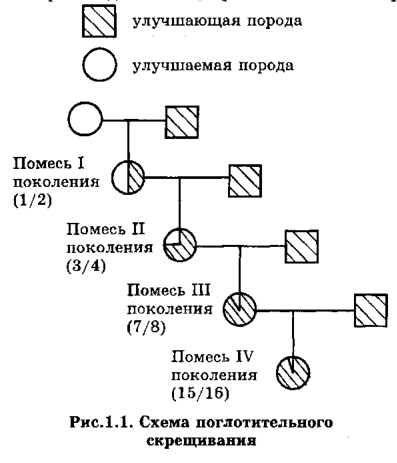
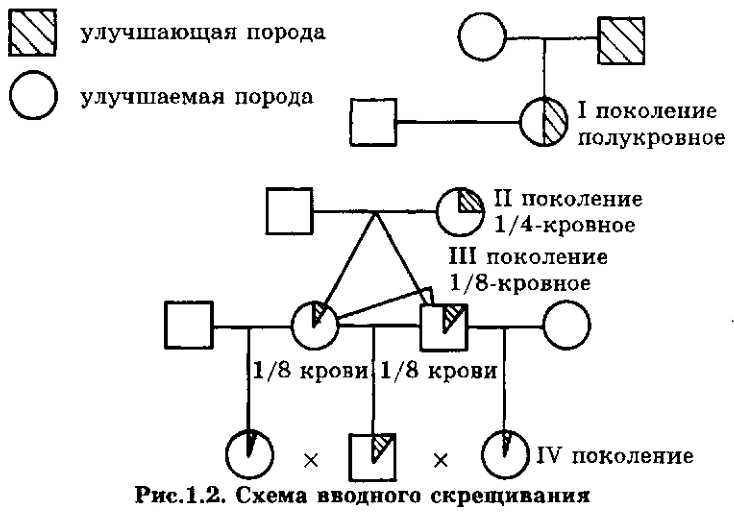
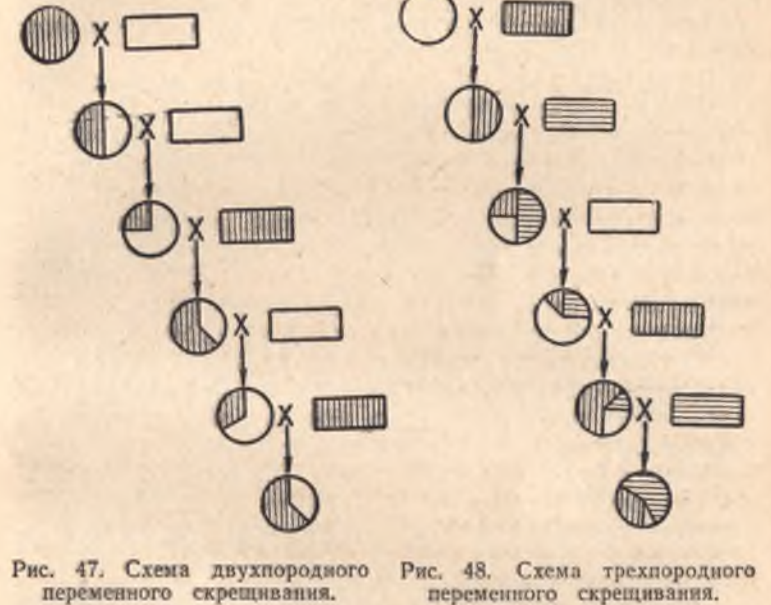
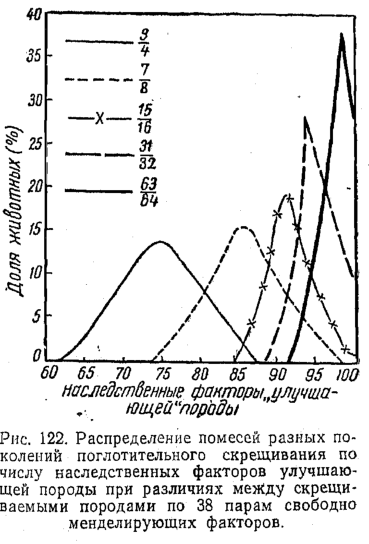
И, теперь, собственно, для начала, пройдёмся по средневековой ошибочной теории «"чистоты" крови» и схемам, демонстрирующим её «наследование», применительно к рис. 37 из книги В.З.Тарантула...
Следуя этой ошибочной теории, мышка-то, очень много и тесно «пообщалась» с человеком... На одинаковые Y, современным адептам ошибочной теории «"чистоты" крови», лучше даже не смотреть!

Продолжение следует...